Let Food Be Thy Medicine I was looking for something different eat that is quick, healthy, hypoallergenic and has all the nutrients my body needs to be healthy, while also support healing the leaky gut. So I pulled out my famous gut-friendly pumpkin banana muffins. This is delicious with a spoon of pumpkin seed butter and [...]
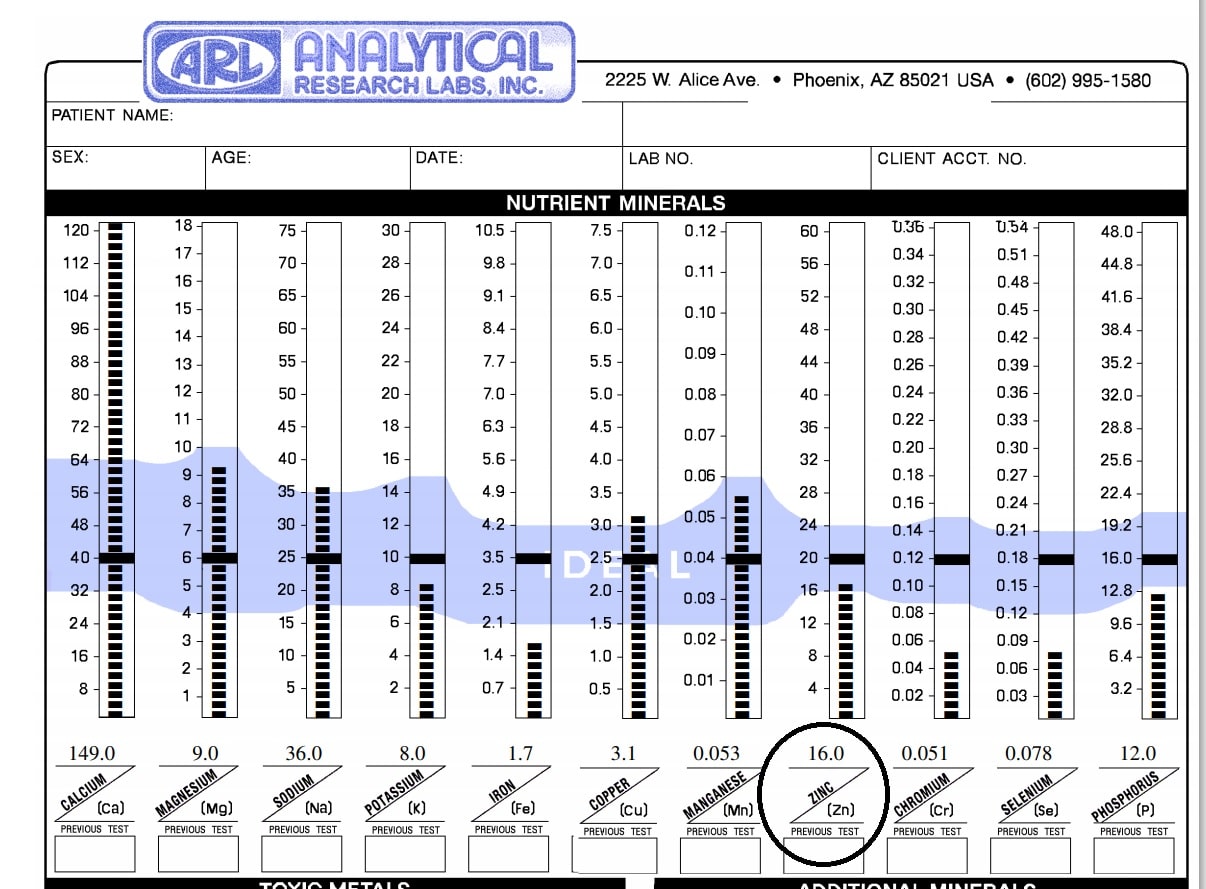
Zinc is a very interesting mineral. It is also one I commonly see low in hair tissue mineral analysis (HTMA). It plays an important role in facilitating hundreds of biochemical reactions. Due to its role in enzymatic function, it can impact metabolic pathways such as carbohydrate, protein, nucleic acid, and lipid metabolism. It is also [...]


")