


Balancing Act: Understanding Copper’s Role in Hormonal Health and Fertility
In the intricate dance of human biology, trace elements play crucial yet often understated roles. Among them, copper stands out for its significant impact on reproductive health. This essential nutrient, though required in minute quantities, plays a pivotal role in various biological processes critical to reproductive function. In this blog, we will dive into the [...]
Unveiling the Hidden World Inside Us: The Intricate Dance of Diet, Metabolism, and Oxalate Production
The topic of oxalate accumulation from citrate by Aspergillus Niger, and specifically the biosynthesis of oxalate from its ultimate precursor, is an area of interest in microbiology and biochemistry. Aspergillus Niger is a common species of fungus known for its industrial applications and its role in various biochemical processes. Here's a summary based on the [...]
Unlocking the Mystery: How Everyday Foods Influence Bladder Health and Oxalate Metabolism
In my clinical practice, I've observed a fascinating pattern: patients grappling with high levels of oxalates often exhibit elevated benzoic acid levels, coupled with diminished hippuric acid. This correlation is particularly notable among those suffering from interstitial cystitis, a chronic condition characterized by bladder pain and discomfort. Understanding the interplay of these compounds in our [...]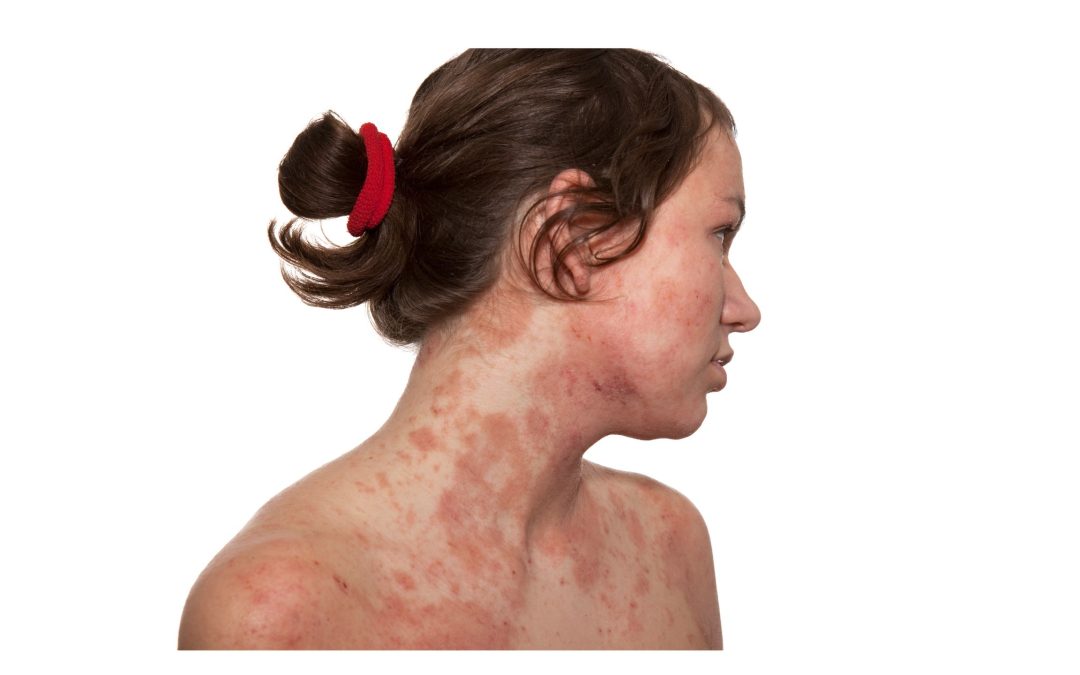
")